DIVISIÓN CELULAR. GUIÓN
(9 sesiones)
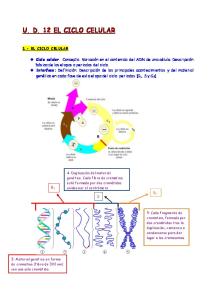
DIVISIÓN CELULAR. - EL CICLO CELULAR. Fases del ciclo celular. Fenómenos generales de cada fase.
- CROMOSOMAS Morfología general de los cromosomas mitóticos Cariotipo y bandeo cromosómico.
- MITOSIS. Descripción de las distintas fases. Movimientos cromosómicos. Huso acromático.
- CITOCINESIS EN CÉLULAS ANIMALES Y VEGETALES. - MEIOSIS. Descripción de las distintas fases. Fenómenos asociados (recombinación, reducción del número de cromosomas.....) Significado biológico de la meiosis.
- CICLOS BIOLÓGICOS. Reducción gamética o terminal (individuos diplontes). Reducción cigótica o inicial (individuos haplontes). Reducción espórica o intermedia (individuos diplohaplontes).
2.- DIVISIÓN CELULAR.En los individuos pluricelulares, las células tienen una existencia mucho más corta que la del organismo entero, por ello, es necesario que las células sean capaces de reproducirse para compensar esa pérdida regular, y que las células hijas posean la misma estructura y los mismos caracteres que la célula madre.
EL CICLO CELULAR. El proceso de la división celular consiste en dos procesos esenciales: la división nuclear (denominada MITOSIS o FASE M) y la división citoplasmática (denominada CITOCINESIS). Pero antes de que una célula pueda dividirse, debe duplicar su masa y todos los elementos que contiene. Sólo de esta forma es posible que las dos nuevas células hijas contengan todos los componentes que necesitan para iniciar su propio ciclo de crecimiento celular, después de la división. La mayor parte del trabajo necesario para la preparación de la división celular se produce de manera invisible durante la etapa de crecimiento del ciclo celular que recibe el nombre de INTERFASE (Gráfico 1). La mayoría de los componentes celulares se forman continuamente a lo largo de todo el período interfásico, sin que pueda establecerse en qué momento concreto se produce cada uno de ellos. Sin embargo, la síntesis de ADN (replicación) constituye una excepción, ya que ocurre
1
durante una etapa limitada y muy concreta de la interfase. Este período se conoce como FASE "S" (S = síntesis) del ciclo celular. El período comprendido entre el final de la última mitosis y el comienzo de la síntesis de ADN, recibe el nombre de FASE G1 (G = gap, en inglés separación) y el período comprendido entre el final de la síntesis de ADN y la siguiente mitosis, recibe el nombre de FASE G2.
FENÓMENOS GENERALES EN CADA FASE: A) INTERFASE. El período G1 es una etapa en la que los genes se transcriben y traducen, sintetizando aquellos enzimas necesarios para hacer funcionar toda la "maquinaria" metabólica celular. Parece ser, que en este período se sintetiza una proteína especial denominada "proteína U" (del inglés "unstable protein", proteína inestable). Cuando su concentración alcanza un valor crítico, desencadena una serie de procesos que inician la duplicación del DNA (Gráfico 2). En el período S, la duplicación del material genético empieza simultáneamente por numerosos puntos a lo largo de la molécula de ADN. En estas zonas, las dos hebras complementarias se separan y cada una de ellas actúa de molde para la síntesis de la nueva cadena. Al final, uniéndose las diferentes secuencias neosintetizadas se tendrá la nueva cadena de ADN completa. La manifestación citológica de la duplicación son las horquillas de replicación. Aparecen por parejas en determinados puntos conocidos como punto de origen y se mueven en sentidos opuestos formando una estructura conocida como burbuja de replicación. Hasta 80 orígenes pueden activarse simultáneamente, constituyendo lo que se conoce como una unidad de replicación. La replicación es un proceso complicado, que se consigue mediante un conjunto de proteínas, entre las que destaca la ADN polimerasa. Cuando la DNA polimerasa empieza la replicación de un trozo de ADN, siempre se sitúa en el extremo 3´ de éste. A continuación va seleccionando, uno a uno, aquellos nucleótidos que tiene libres por su entorno, apareándolos con la hebra patrón. Uniendo, cada nucleótido al adyacente mediante un enlace covalente, se va sintetizando la nueva cadena, la cual va creciendo en dirección 5´───> 3´. Una vez que la información genética es duplicada en su totalidad, es decir, se llegan a formar dos dobles hélices de ADN, cada una de ellas complementaria en cuanto a secuencia de nucleótidos a la que sirvió de patrón, termina el proceso de replicación. Parece ser que las histonas viejas permanecen unidas en nucleosomas sobre una hélice hija de ADN, mientras que sobre la otra hélice se unen nuevas histonas que forman nuevos nucleosomas. Terminada la replicación del material genético, la célula contiene doble cantidad de ADN que una célula normal. Este dato nos revela que estamos en el tercer período de la interfase: la fase G2 (Gráfico 3). Durante este período se sintetizan determinadas proteínas necesarias en la mitosis. Una de estas proteínas es la enzima proteinquinasa, la cual sería la responsable de: la desintegración de la envoltura nuclear en las primeras etapas de la e intervendría en la fosforilación de las histonas H1, lo que causaría la cromosómica.
2
fase M.
condensación
B) DIVISIÓN
CELULAR.
Al iniciarse la división celular la cromatina se va concentrando. Como la cromatina de las células está fraccionada en trozos, cuando ésta se condensa se constituyen unos cuerpos independientes (uno por cada fragmento de cromatina), conocidos como cromosomas. Estos, durante la mitosis, se alinearán en el centro de la célula y se separarán hacia los extremos opuestos de la célula. Después, el citoplasma se ha de segmentar de tal manera que asegure que cada célula hija reciba no sólo un conjunto completo de cromosomas, sino también los elementos y los orgánulos citoplasmáticos necesarios. Aunque estos procesos, la división nuclear (mitosis) y la división citoplasmática (citocinesis) son experimentalmente separables, generalmente se producen en estrecha sucesión, de forma que la citocinesis se inicia hacia el final de la mitosis.
CROMOSOMAS. A) MORFOLOGÍA DE LOS CROMOSOMAS.- En las primeras etapas de la mitosis, cada cromosoma está formado por 2 subunidades iguales, conteniendo cada una de ellas una sóla molécula de ADN. Cada una de estas estructuras recibe el nombre de CROMÁTIDA. Cada cromátida está constituida por una de las dos copias de ADN que se han formado por replicación en la fase "S"; por lo tanto, las cromátidas y las moléculas de ADN que las constituyen son idénticas. El motivo por el que las dos copias de ADN permanecen unidas temporalmente es para que el reparto de las mismas sea más fácil.
Aclaración.- Los cromosomas homólogos regulan los mismos caracteres hereditarios, aunque cada uno de los cromosomas del par, puede determinar una modalidad del carácter distinta. Cada cromosoma homólogo deriva de un progenitor (padre y madre). Las cromátidas regulan los mismos caracteres hereditarios y además como su ADN es idéntico, determinan la misma modalidad del carácter. Se puede decir que las cromátidas son idénticas y los cromosomas homólogos similares . Las dos cromátidas de un cromosoma se mantienen unidas temporalmente, por un punto conocido como CENTRÓMERO. Puede utilizarse esta zona, para dividir cada cromátida en dos partes llamadas brazos, las cuales pueden ser iguales o diferentes. Cada cromátida presenta a nivel del centrómero, una subestructura discoidal denominada
CINETÓCORO, que actúa como centro organizador de microtúbulos. A partir de este centro se forman los filamentos tubulares que dirigen los movimientos cromosómicos durante la mitosis y la meiosis.
Las cromátidas presentan zonas estrechas conocidas como constricciones. La más evidente se encuentra situada a nivel del centrómero y se conoce como constricción primaria. Pueden existir otras constricciones, que se conocen como secundarias, que por presentarse siempre en la misma situación en los cromosomas resultan muy útiles para su identificación. Los extremos distales de cada brazo cromosómico se denominan TELÓMEROS. La cromatina de esta zona, durante la interfase, se mantiene en contacto con la membrana nuclear para mantener así el orden nuclear. Ciertos cromosomas presentan unas formaciones, redondas o alargadas, conocidas como
3
SATÉLITES, que están unidas a cada cromátida por un delgado filamento cromático. Su diámetro puede ser igual al de la cromátida o ser más pequeño. El filamento de unión puede ser largo o corto. Como el satélite y el filamento son constantes en forma y tamaño para el cromosoma, pueden servir para identificar al cromosoma (ejemplo: cromosomas 13, 14, 15 y 21 del cariotipo humano). b) TIPOS DE CROMOSOMAS.- Cada cromosoma puede ser identificado por su forma y tamaño. Su forma viene determinada por la posición del centrómero, que, a su vez, condiciona el lugar de anclaje a las fibras del huso mitótico. Atendiendo a las distintas posiciones del centrómero, los cromosomas se clasifican en los siguientes 4 tipos: TELOCÉNTRICOS: Presentan el centrómero en el extremo, de manera que cada cromátida está constituida por un sólo brazo. ACROCÉNTRICOS: Con el centrómero dispuesto de tal manera que el cromosoma queda dividido en un brazo muy corto y otro muy largo. SUBMETACÉNTRICOS: la posición del centrómero determina que la longitud de los brazos es desigual. En anafase adquieren la forma de "L". METACÉNTRICOS: Cuando la posición del centrómero es medial y, por tanto, los dos brazos resultantes son iguales. C) CARIOTIPO.- Se denomina cariotipo al conjunto de cromosomas característicos de una especie. En el caso de los humanos, las parejas de cromosomas son numeradas del 1 al 22, siendo conocidos todos estos ellos como autosomas. Dos cromosomas que no presentan mucho parecido son etiquetados como "X" e "Y"; se trata de los cromosomas sexuales. Los cromosomas de cada par son idénticos desde el punto de vista morfológico y genéticamente son similares ya que contienen información para los mismos caracteres, se denominan cromosomas homólogos. De cada pareja de homólogos, uno de los cromosomas deriva de la dotación paterna que fue otorgada en el espermatozoide y el otro procede de la materna. Todas las células de un individuo, excepto las sexuales, y todos los individuos de una misma especie, poseen el mismo número de cromosomas. Se dice que ésta es su dotación diploide y puede representarse como "2n". Este número diploide puede ser distinto para diferentes especies, así, por ejemplo: Hombre "2n" = 46 Gato "2n" = 38 Ratón "2n" = 40 Perro "2n" = 78
Chimpancé "2n" = 48 Patata "2n" = 48 Tabaco "2n" = 48 Caballo "2n" = 64
Los gametos o células sexuales (óvulos, espermatozoides, esporas de helecho, ciertas algas......) son haploides, es decir, contienen un sólo juego cromosómico. Este número de cromosomas puede representarse como "n". Hombre "n" = 23
4
Hay organismos, como el trigo, que contienen más de dos juegos cromosómicos y se denominan triploides (3n), tetraploides (4n) y, en general, poliploides. D)BANDEO CROMOSÓMICO.- Cuando los cromosomas se tiñen con determinados colorantes como la fucsina en la técnica de Feulgen, aparecen unas bandas más oscuras (heterocromatina) intercaladas entre otras claras (Eucromatina) (Gráfico 4). La Heterocromatina está constituida por ADN en estado muy condensado, el cual permanece en este estado incluso durante la interfase, cuando las otras partes del cromosoma han adoptado una forma más dispersa y constituyen lo que se conoce como cromatina. El DNA de la heterocromatina no se transcribe a ARN, por lo que se dice que es inactivo (parece ser que la extraordinaria condensación de esta cromatina impide la expresión de sus genes). La Eucromatina corresponde a las zonas del DNA que se transcriben, por lo tanto, es en ella donde se encuentran los genes. Como las bandas oscuras y claras se pueden reconocer por su grosor y separación, se las ha numerado empezando por el centrómero y siguiendo por ambos brazos de cada cromosoma. De esta forma, disponemos de un sistema cartográfico (mapa) del cromosoma que nos permite referirnos a cualquier punto de él. Por ejemplo, las siglas Xq28 designan la octava subbanda de la banda 2 del brazo largo del cromosoma X. Utilizando este sistema, en la actualidad se están realizando una serie de estudios para localizar los genes dentro de los cromosomas.
5
MITOSIS. La MITOSIS es el proceso de división celular, por el cual, surgen células con el mismo número de cromosomas que la célula madre. Tradicionalmente, la mitosis se estructura en cuatro etapas:
PROFASE .- Condensación de cromatina y aparición de cromosomas. METAFASE.- Los cromosomas se ordenan en el centro de la célula. Huso mitótico.
ANAFASE.- Los cromosomas se parten a nivel de sus centrómeros y las dos cromátidas se desplazan hacia polos opuestos de la célula.
TELOFASE.- Separación citoplasmática y formación de dos células hermanas. (Regla nemotécnica: Promete a Ana un teléfono) (Gráfica 9).
DESCRIPCIÓN DE LAS DISTINTAS FASES: A) PROFASE (pro = delante, antes). La transición desde la fase G2 a la profase no es un suceso estrictamente definido. La primera evidencia de que se ha pasado al estado de profase es que la doble membrana nuclear empieza a fragmentase en vesículas. Las fibras de cromatina, que previamente se habían duplicado en la fase "S", se condensan lentamente formando cromosomas bien definidos. Cada cromosoma está formado por 2 cromátidas hermanas unidas por un punto llamado centrómero. Mientras los cromosomas se condensan, el nucléolo empieza a descondensarse y desaparece progresivamente. Toda la maquinaria de transcripción del nucléolo se empaqueta y se guarda entre los cromosomas 13, 14, 15, 21 y 22 (en las células humanas). La masa de microtúbulos citoplasmáticos que forman parte del citoesqueleto se disgrega formando una gran reserva de moléculas de tubulina. Estas moléculas, se supone que se utilizan posteriormente en la construcción del huso mitótico. En las CÉLULAS ANIMALES, el par de centríolos se duplican originando dos diplosomas diferentes. Cada diplosoma organiza a los microtúbulos del entorno, constituyendo una estructura radial conocida como áster (áster = estrella). Al comenzar la profase, los dos centrosomas con sus respectivos ásteres se hallan situados uno junto al otro, pero a lo largo de este período se irán separando hasta colocarse en polos opuestos de la célula. De esta manera y al alargarse las fibras de los ásteres, se forma el huso mitótico. Las CÉLULAS VEGETALES carecen de centríolos y los filamentos del huso parten de dos zonas más densas del citoplasma.
6
B)
METAFASE (meta = reunidos).
La metafase se inicia cuando la envoltura nuclear está totalmente desintegrada, de manera que las estructuras del citoplasma pueden mezclarse con las del nucleoplasma. En cada cromosoma, cerca de los centrómeros, se desarrollan unas estructuras denominadas cinetocoros. Estas formaciones están unidas a un conjunto de microtúbulos que se denominan como fibras cinetocóricas. Estas fibras crecen desde los cinetocoros y avanzan en sentidos opuestos hasta que entran en interacción en cada polo, con las fibras del huso. Parece ser, que son estas fibras las responsables de la alineación de los cromosomas a medio camino entre los dos polos del huso, manteniéndolos en tensión en el plano ecuatorial o placa metafásica.
C) ANAFASE (aná = atrás). La anafase empieza bruscamente cuando los cinetocoros de cada cromosoma se separan, y a medida que se acortan las fibras cinetocóricas , cada cromátida de un par es arrastrada
hacia un polo opuesto.
La separación de las cromátidas es el resultado de las tensiones generadas por las fibras cinetocóricas, las cuales se deslizan hacia los polos y ejercen una fuerza de tracción sobre las cromátidas. Esta tensión aplicada sobre los cromosomas causa la rotura de sus centrómeros, aunque se sospecha que en este proceso también participan determinados enzimas. Desde el momento en que se separan las cromátidas hermanas, éstas pasan a ser conocidas como cromosomas hijos o cromosomas anafásicos. D) TELOFASE. La telofase comienza cuando los cromosomas hijos se han separado por completo en dos grupos iguales, uno en cada polo del huso. Llegado este momento, las fibras cinetocóricas desaparecen. Alrededor de cada grupo de cromosomas, comienza a formarse las membranas nucleares interna y externa, que se continúan con la membrana del retículo endoplasmático. También se forman la lámina nuclear y los poros nucleares.
Reaparece el nucléolo y los cromosomas se vuelven a descondensar, retornando al
estado de cromatina.
7
CITOCINESIS EN CÉLULAS ANIMALES Y VEGETALES. El objetivo de la citocinesis es dividir el citoplasma, al mismo tiempo que se reparten entre las dos células hijas, los orgánulos presentes en él. El proceso difiere, según se trate de células animales o vegetales, por ello estudiaremos la citocinesis en cada caso.
En las CÉLULAS ANIMALES, la citocinesis suele iniciarse en la anafase, apareciendo como primer signo de la misma, una pequeña invaginación de la membrana plasmática a lo largo del plano de la placa metafásica. Poco a poco, esta invaginación se hará más profunda, provocando la estrangulación celular. Parece ser que este surco de segmentación lo origina un anillo de fibras de actina y miosina, situado entre los dos núcleos, que se estrecha progresivamente como si se tratara de un lazo corredizo, haciendo que el surco de segmentación sea cada vez más profundo, hasta que se separan por completo las dos células hijas (Gráfica 12).
En las CÉLULAS VEGETALES, la pared celular rígida que las rodea no permite el estrangulamiento y la citocinesis ocurre por formación de un tabique de separación entre las dos células hijas. La nueva pared celulósica, denominada fragmoplasto, se forma por la fusión de vesículas procedentes del aparato de Golgi que cargadas de celulosa y otros polisacáridos, se alinean en el plano ecuatorial y luego se fusionan. Entre las dos células hijas y atravesando el fragmoplasto, subsisten finos puentes citoplasmáticos: los plasmodesmos. Cuando los extremos de las membranas del fragmoplasto terminan por fusionarse con la membrana celular, queda un espacio vacío entre ellas en el que se deposita pectina, la cual constituirá la lamina media (Gráficas 13 y 15). Las células vegetales no pueden reorganizarse después de divididas debido a las rígidas paredes celulares que las rodean, por ello, ya que el crecimiento de una planta tiene que estar organizado, las células vegetales tienen que tener controlada la división antes de que comience. Los microtúbulos desempeñan un papel crucial en este sentido, ya que determinan el lugar en el que se acumularán las vesículas cargadas de celulosa y por tanto determinarán el plano de división celular y de crecimiento del órgano (Gráfica 14).
8
MEIOSIS. SIGNIFICADO BIOLÓGICO DE LA MEIOSIS.- Las células sexuales (gametos) son haploides, debido a que son producidas por medio de una forma especial de división celular: la meiosis. Los individuos pluricelulares se reproducen gracias a un mecanismo particular conocido como fecundación, que consiste en la unión de dos células especiales llamadas gametos. Cada gameto procede de un individuo de sexo distinto y tras la unión de ambos se forma una célula llamada huevo o cigoto que es el punto de partida del desarrollo embrionario. Si las células que constituyen los espermatozoides y los óvulos fueran células corrientes, tendrían el mismo número de cromosomas que las restantes. Cuando en la fecundación se unieran, constituirían un huevo o cigoto con el doble número de cromosomas, el cual al desarrollarse produciría un descendiente de células con doble número de cromosomas. El resultado sería que el número de cromosomas se duplicaría en cada generación. Para evitar esto, que sin duda sería un problema, el organismo produce a los gametos mediante una modalidad especial de división celular: la MEIOSIS . Este término deriva del griego y significa "disminución", queriendo resaltar así que la finalidad principal de este proceso es precisamente,
disminuir a la mitad el número de cromosomas de estas células.
La comprensión de este proceso se obtuvo como resultado de unas observaciones que se hicieron en 1883 sobre un gusano (Parascaris equorum). Se encontró que los núcleos del óvulo y del espermatozoide sólo contenían 2 cromosomas cada uno, mientras que el huevo fecundado contiene 4 cromosomas. A partir de estos datos se derivan 2 conclusiones: - Que el carácter de la progenie viene determinado por una contribución materna y paterna, que son iguales. - Y que las células sexuales deben formarse por medio de un tipo especial de división nuclear, en el que la dotación cromosómica queda dividida exactamente por la mitad.
La meiosis consta de dos divisiones consecutivas: Antes de la primera división, los cromosomas se duplican durante el período "S" de la interfase, estando formados cada uno de ellos por un par de cromátidas hermanas. En la PRIMERA DIVISIÓN MEIÓTICA se separan los cromosomas homólogos y el número cromosómico diploide es reducido a haploide, esto es, cada célula hija contiene uno sólo de los 22 autosomas, y uno de los cromosomas sexuales. Los cromosomas homólogos, antes de la separación son capaces de reconocerse mutuamente y quedan físicamente unidos durante un cierto tiempo. Es en este estado de apareamiento cromosómico, cuando ocurre uno de los fenómenos más importantes de la meiosis: el entrecruzamiento cromosómico que trae como consecuencia la recombinación genética. Entre la primera y la segunda división meiótica NO existe fase "S". En la SEGUNDA DIVISIÓN MEIÓTICA se separan las cromátidas que constituyen cada uno de los cromosomas segregados, de manera que las células que surgen conservan el número haploide de cromosomas, pero tienen su contenido de DNA dividido a la mitad de la células diploides G1 de los tejidos somáticos. Conclusión: La meiosis consiste en 2 divisiones celulares sucesivas que se producen tras una única
9
fase de replicación cromosómica, por lo que se producen 4 células haploides a partir de cada célula que sufre la meiosis. ∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙∙
DESCRIPCIÓN DE LAS DISTINTAS FASES. Antes de que ocurran las divisiones meióticas, se duplica el DNA durante la interfase, y cada cromátida da lugar a su cromátida gemela, quedando unidas por el centrómero. A partir de este momento transcurren 2 divisiones sucesivas, cada una de las cuales subdividida en 4 etapas, como en la mitosis.
A) PRIMERA
DIVISIÓN MEIÓTICA.
PROFASE I.- Es la etapa que tarda más tiempo en transcurrir y en la que se dan los acontecimientos más importantes de la meiosis. Los dos acontecimientos más característicos de esta etapa son: que la envoltura nuclear permanece intacta, desapareciendo sólo al final se desintegra el nucléolo. y que durante ella, se constituye el huso, a partir de los microtúbulos
cuando también
citoplasmáticos.
La profase I de la meiosis es tan compleja que se subdivide en 5 etapas secuenciales: Leptoteno, zigoteno, paquiteno, diploteno y diacinesis.
LEPTOTENO (gr. Leptós = delgado, fino).
Esta etapa comienza cuando la cromatina interfásica se condensa y aparece cada uno de los cromosomas celulares. Los cromosomas tienen sus cromátidas tan estrechamente unidas que aparecen como finas y largas hebras cuyos extremos se encuentran unidos a la envoltura nuclear .
ZIGOTENO (gr. dsygón = yugo, yunta). Esta etapa comienza con el apareamiento de los cromosomas homólogos, fenómeno que también es conocido con el nombre de SINAPSIS. Los pares de cromosomas se unen por completo e incluso se cree que cada gen queda yuxtapuesto a su gen homólogo del cromosoma opuesto. Los cromosoma "X" e "Y" no se sinapsan por completo. Puesto que las cromátidas de cada cromosoma todavía no se distinguen, la estructura resultante de este apareamiento parece estar formada solamente por dos elementos, de ahí que se le de el nombre de complejo bivalente.
10
PAQUITENO (gr. pach_s = denso, grueso, espeso). En esta etapa tiene lugar el ENTRECRUZAMIENTO CROMOSÓMICO, nombre con el que se conoce al fenómeno de intercambiar fragmentos de DNA entre cromosomas homólogos. Este entrecruzamiento implica: la rotura de un trozo de ADN de doble hélice perteneciente a una cromátida y la del trozo equivalente de la cromátida del cromosoma homólogo. el intercambio de los fragmentos originados y su unión recíproca. Esto supone que aquel fragmento originado en el cromosoma paterno se insertará junto con los genes del cromosoma materno y viceversa. La consecuencia es que los cromosomas que surgen tras esta etapa son considerados como diferentes a los originales, ya que la secuencia de genes (o en última instancia del ADN que los forma) es distinta. Se dice que cada cromosoma está constituido por una nueva combinación genética, es decir se ha producido una RECOMBINACIÓN GENÉTICA entre los cromosomas homólogos.
DIPLOTENO (gr. diplos = doble). Aquí, los cromosomas homólogos se repelen, aunque permanecen unidos en ciertos puntos llamados quiasmas (gr. chiasma = cruz). Estas zonas reflejan los lugares en donde se ha producido entrecruzamiento. Se suele decir que el quiasma es la manifestación citológica del entrecruzamiento, y la consecuencia genética del mismo es la recombinación genética. En esta etapa, los cromosomas pueden mostrar una intensa actividad de síntesis de ARN (transcripción). Esta fase puede durar desde unos meses a inclusos años. Por ejemplo, en la mujer, los ovocitos se forman en el quinto mes de vida fetal y permanecen es esta fase hasta que la ovogénesis continua a partir de la pubertad.
DIACINESIS (gr. dia = a través y Kinesis = movimiento) En esta fase cesa la síntesis de ARN y los cromosomas se condensan y engruesan, iniciando un desplazamiento que los situará en la placa ecuatorial. Se observa claramente que cada bivalente está formado por 4 cromátidas separadas, de ahí que al conjunto se le conozca ahora como tétrada. Cada par de cromátidas hermanas están unidas por sus centrómeros, mientras que las cromátidas no hermanas que se han entrecruzado están unidas por quiasmas.
11
Nombre ESTRUCTURA en período PAQUITENO
Nombre ESTRUCTURA en
DIACINESIS.
COMPLEJO BIVALENTE ──────────────────────────── TÉTRADA
PROCESO CITOLÓGICA Apareamiento cromosomas homólogos
ESTRUCTURA Complejo bivalente
OBSERVACIÓN Quiasma
CONSECUENCIA
FORMADA
Recombinación genética
FINAL PROFASE I.- Al final de la diacinesis comienza la desaparición de la envoltura nuclear y del nucléolo. Se forma el huso entre los polos celulares, al mismo tiempo que empiezan a formarse los microtúbulos cinetocóricos que colocarán a los cromosomas en el plano ecuatorial. La METAFASE I, ANAFASE I y TELOFASE I, transcurren como en la mitosis. La única diferencia que presenta la meiosis con respecto a aquella, es que en esta primera división celular cada célula hija ha recibido un cromosoma de cada par de homólogos, es decir, se han segregado cromátidas hermanas unidas por un centrómero, en lugar de cromátidas separadas como en la mitosis (Gráfica 11). Tras una INTERFASE división meiótica.
B)
II transitoria (sin replicación de ADN), se comienza con la segunda
SEGUNDA DIVISIÓN MEIÓTICA.
Su estudio se divide en 4 etapas conocidas como PROFASE II, METAFASE II, ANAFASE II y TELOFASE II. Los fenómenos que se desarrollan en ellas son semejantes a los de la mitosis, la única diferencia es que se parte de células haploides. Cada cromosoma, constituido por dos cromátidas se separará, originando células haploides, con cromosomas constituidos por una sola copia de ADN.
12
Conclusión: La meiosis consiste en dos divisiones celulares sucesivas que se producen tras una única fase de replicación cromosómica, por lo que se producen 4 células haploides a partir de cada célula que sufre la meiosis.
INTERFASE
│ Período "S" (replicación del ADN).
1ª DIVISIÓN MEIÓTICA │ PROFASE I Fase Leptoteno │ " Zigoteno │ " Paquiteno │ " Diploteno │ " Diacinesis │ METAFASE I │ ANAFASE I │ TELOFASE I 2ª DIVISIÓN MEIÓTICA │ PROFASE II │ METAFASE II │ ANAFASE II │ TELOFASE II
CICLOS BIOLÓGICOS Se entiende por ciclo biológico, al conjunto de etapas por las que transcurre un organismo, desde que se origina como cigoto (por fecundación de los gametos), hasta que él mismo alcanza la edad adulta y produce nuevos gametos. A lo largo del ciclo se pasa por una etapa diploide y otra haploide, que son distintas en los diferentes organismos según el momento en el que tenga lugar la meiosis. Se distinguen 3 tipos de ciclos:
CICLO BIOLÓGICO CON REDUCCIÓN GAMÉTICA o TERMINAL (INDIVIDUOS DIPLONTES). La meiosis tiene lugar en la fase previa a la formación de los gametos, que son haploides. Tras la fecundación se forma un cigoto diploide, que se desarrolla y origina un adulto diploide. Se da en los Metazoos y en algunos Protistas.
13
CICLO BIOLÓGICO CON REDUCCIÓN CIGÓTICA o INICIAL (INDIVIDUOS HAPLONTES). Sólo se forman gametos cuando las condiciones desfavorables del medio, así lo aconsejan. Tras la fecundación, se forma un cigoto diploide que inmediatamente se encapsula, originando una espora de resistencia capaz de aguantar mientras las condiciones no mejoren. Cuando las condiciones ambientales son favorables, el cigoto experimenta la meiosis y da lugar a 4 individuos adultos haploides. Estos pueden reproducirse vegetativamente, por divisiones celulares o por fragmentación. Cuando las condiciones entorno, de nuevo empeoran, producen gametos y se inicia de nuevo el ciclo. Se da en determinadas algas unicelulares, como Chlamydomonas.
CICLO BIOLÓGICO CON REDUCCIÓN ESPÓRICA o INTERMEDIA ( INDIVIDUOS DIPLOHAPLONTES). A lo largo del ciclo vital de estos organismos, se distinguen 2 formas de individuos, que van alternando generacionalmente; es decir, unos son diploides y los de la generación siguiente son haploides. Estudiaremos el caso de los helechos. La fase adulta es diploide y es conocida como ESPOROFITO. En los esporangios, se forman las esporas por meiosis, de manera que estas son haploides. Las esporas son liberadas y si alcanzan algún lugar favorable, germinan originando individuos haploides denominados GAMETOFITOS. Estos, desarrollan las estructuras que formarán los gametos, también haploides. La fecundación del gameto femenino por el masculino da lugar a un cigoto diploide, que al desarrollarse forma un nuevo individuo adulto como el del principio: un nuevo ESPOROFITO
AMPLIACIÓN. 2.2. TIPOS DE CROMATINA. En el núcleo interfásico, se pueden apreciar dos estados de condensación de la cromatina: Hay zonas donde el empaquetamiento es menos denso (los bucles están distendidos) y se conoce como eucromatina, mientras que en otras zonas, está empaquetada en una configuración altamente densa y entonces se conoce como heterocromatina. En aquellas zonas donde la cromatina está poco condensada, los bucles se encuentran distendidos y la RNA polimerasa y el resto de enzimas que participan en la TRANSCRIPCIÓN pueden acceder hasta el DNA. Por lo tanto, en la eucromatina se produce la transcripción. En las zonas de heterocromatina, debido al mayor grado de empaquetamiento, el ADN no se transcribe y por lo tanto permanece funcionalmente inactivo. Existen 2 clases de heterocromatina: La H. constitutiva. Son zonas que siempre se encuentran condensadas y por lo tanto su ADN nunca se transcribe, desconociéndose su función y su estructura. y la H. facultativa. Se trata de otras zonas que permanecen inactivas desde el momento de la diferenciación celular. Parece ser que a medida que se desarrollan las células, aquellos genes cuya información ya no es necesaria se inactivan empaquetandose permanentemente en forma condensada.
14
2.3. IMPORTANCIA DE LAS PROTEÍNAS NO HISTÓNICAS. El ADN de todos los organismos está íntimamente asociado con una amplia variedad de proteínas. Tradicionalmente, las proteínas que se unen al ADN de los eucariotas se clasifican en dos grupos: las histonas, que actúan como elementos estructurales organizando el ADN dentro del núcleo celular. y las proteínas no histonas, que actúan con funciones muy diversas. Por ejemplo, algunas actúan como enzimas en procesos tan importantes como la síntesis de ADN. Entre las proteínas no histonas, se cuentan aquellas que se unen a secuencias específicas de ADN, controlando la transcripción del mismo. Este control se demostró, en primer lugar, en bacterias de E. coli, al observar que hay una proteína REPRESORA que se une al ADN, de modo que inhibe la síntesis de aquellos ARNs capaces de sintetizar enzimas del metabolismos de la lactosa. Esta proteína represora se une a una secuencia específica de ADN, conocida como OPERADOR, superponiendose en parte a una región adyacente en la que se une la RNA polimerasa al DNA, conocida como región PROMOTORA. Cuando la proteína represora está unida al ADN, bloquea el acceso de la ARN polimerasa a su región de unión, impidiendo así la transcripción del ADN adyacente. Cuando existe lactosa que metabolizar dentro de la célula, se sintetiza una pequeña molécula, llamada alolactosa, que uniéndose a la proteína represora hace que esta se desenganche del ADN, permitiendo que se inicie la transcripción. Se dice entonces que el gen está desreprimido. Sobre este gen se sintetiza ARNm, que por la traducción producirá una enzima que a su vez actuará hidrolizando la lactosa. De esta forma, en la célula solo se producen enzimas hidrolíticos de la lactosa cuando hay moléculas de ésta disponibles.
NIVELES DE ORGANIZACIÓN DE LOS CROMOSOMAS. El DNA de todos los organismos está íntimamente asociado con una amplia variedad de proteínas. Un grupo lo constituyen las proteínas histonas, las cuales organizan el ADN dentro del núcleo celular. Otras, las proteínas no histonas, son enzimas que actúan en diferentes procesos como, por ejemplo, llevar a cabo la duplicación del ADN (las DNA-polimerasas). Existen diferentes tipos de proteínas histonas. Las conocidas como H2A, H2B, H3 y H4, constituyen unos núcleos proteicos alrededor de los cuales se enrolla el ADN de doble hebra. Esta asociación, cuya estructura recuerda a un "collar de perlas" o "cuentas ensartadas" es lo que se aprecia al microscopio como una fibra cromatínica de 30 nm. La fibra cromatínica de 30 nm se pliega en forma de grandes bucles, los cuales pueden llegar a compactarse en extremo durante la mitosis, constituyendo el cromosoma. Estos son los diferentes grados de empaquetamiento que presenta la molécula de ADN dentro del núcleo, desde la más elemental que es cada una de las "cuentas" del "collar de perlas" (considerada como la unidad de organización fundamental y conocida como nucleosoma) hasta la cromatina, que es el aspecto con el que se observa el ADN durante la interfase o los cromosomas durante la mitosis (Gráficos 6, 7 y 8).
15